پیام رسانی و سازگاری
در حضور مولکول نشانه نیز پاسخ رفته رفته کاهش می یابد. این فرآیند سازگاری نام دارد.
مرکز یادگیری سایت تبیان - مرجان سلیمانیان
پس از حذف یک پیام از محیط، پاسخ سلول به آن باید متوقف شود. در بسیاری موارد، پایان پیام رسانی سریع است. مثلا در سلول های استوانه ای چشم در نبود نور، Gαt به سرعت و با کمک پروتئین های فعال کننده (GAP) GTPase می تواند GTP خود را به GDP هیدرولیز کند و به صورت غیر فعال درآید. بلافاصله آنزیم cGMP فسفو دی استراز که اثر کننده Gαt است، غیر فعال می شود. سپس فعالیت سریع آنزیم گوانیلیل سیکلاز باعث می شود سطح cGMP در سلول به میزان عادی باز گردد و کانال های کاتیونی باز شوند. این کار تضمین می کند که در نبود نور، تولید پیام بینایی از سلول های استوانه ای به سرعت قطع شود.
در موارد دیگر، پاسخ با سرعت کمتری پایان می یابد. مثلا عمل پروتئین کینازها می تواند پس از مدتی در نتیجه عمل پروتئین فسفاتازهای عمومی و اختصاصی جبران شود. در نتیجه با غیر فعال شدن پروتئین کینازها، فسفاتازها پاسخ سلول را متوقف می کنند. پیک های ثانویه نیز می توانند حذف شوند، مثلا cAMP توسط cAMP فسفودی استراز هیدرولیز می شود.
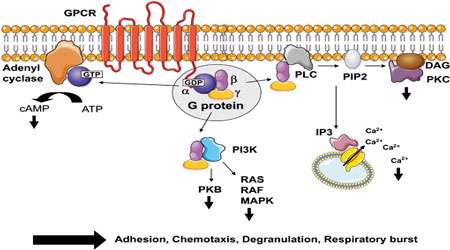
از طرفی حتی در حضور مداوم یک پیام نیز پاسخ سلول ثابت نمی ماند و با زمان کم رنگ می شود. این ویژگی سازگاری نام دارد. سازگاری به پیام ورودی ویژگی تمام مسیرهای پیام رسانی سلولی است، به طوری که سلول بتواند کماکان به تغییر در پیام پاسخ دهد، یعنی در حضور یک آگونیست، پاسخ سلول رفته رفته کم می شود و به سطح ثابتی می رسد که پایین تر از سطح پاسخ اولیه است. این فرآیند بازخوردی منفی به طور عمومی در تمام مسیرها و از جمله در مسیر پیام رسانی با گیرنده های جفت شده با G پروتئین (GPCR)، تیروزین کینازهای گیرنده (RTKها) و گیرنده های سایتوکاین دیده می شود.
به فرآیندهای سازگاری بی حس شدن نیز گفته می شود. بی حس شدن به معنی آن است که سلول مدتی پس از حضور یک آگونیست به آن حساسیت کمتری نشان می دهد. به علاوه سلول این بی حس شدن را تا مدتی پس از حذف آگونیست نیز حفظ می کند و اگر در این مدت آگونیست مجددا به محیط افزوده شود، پاسخ دوم کمتر از پاسخ اول خواهد بود.
بی حس شدن غالبا در سطح گیرنده اتفاق می افتد. یک نمونه از سازگاری یک مسیر GPCR را بررسی می کنیم:
گیرنده: با اتصال اپی نفرین به گیرنده بتا- آدرنرژیک، صورت بندی گیرنده در بخش سیتوپلاسمی تغییر می کند. با تغییر صورت بندی، علاوه بر آغاز پیام رسانی، گیرنده می تواند به پروتئین های دیگری هم متصل شود و سوبسترای برخی آنزیم ها نیز قرار گیرد.
یکی از این آنزیم ها پروتئین کیناز A (PKA) غاست که خود می تواند در پی فعال شدن این گیرنده فعال شود. PKA گیرنده را فسفریله می کند. گیرنده فسفریله کارایی کمی در اتصال به Gαs و فعال کردن آن دارد. یک پروتئین دیگر به نام کیناز گیرنده بتا- آدرنرژیک (BARK) که ویژه این گیرنده است نیز می تواند صورت بندی فعال گیرنده (در اتصال با اپی نفرین) را شناسایی و آن را فسفریله کند.
* اتصال ارستین: پروتئینی به نام ارستین به گیرنده فسفریله متصل می شود و از فعالیت آن جلوگیری می کند.
* اندوسیتوز گیرنده: ارستین متصل شده با فراخوانی کلاترین و پروتئین وفق دهنده AP-2 باعث اندوسیتوز گیرنده در وزیکول های درونی می شود و از تعداد گیرنده ها در سطح سلول می کاهد. گیرنده ها به این ترتیب در درون سلول ذخیره شده اند.
* تجزیه اندوزومی گیرنده: گیرنده های اندوسیتوز شده ممکن است به لیزوزوم ها بپیوندند و تجزیه شوند.
* مهار بیان گیرنده: در حضور دراز مدت پیام، بیان ژن رمز کننده یک گیرنده ممکن است مهار شود. تفاوت اثر PKA و BARK (که هر دو گیرنده را فسفریله می کنند) دو نوع متفاوت بی حس شدن را مشخص می کند.
* PKA می تواند توسط انواعی از GPCR ها فعال شود و سپس انواع آنها را فسفریله (مهار) کند. این بی حس شدن هترولوگ (نا همسان) می باشد که در آن سلول در اثر حضور یک لیگاند نسبت به لیگاندهای دیگر (با گیرنده های متفاوت) نیز بی حس می شود.
* BARK فقط به GPCR از نوع بتا- آدرنرژیک متصل می شود و آن را فسفریله (مهار) می کند. این بی حس شدن مختص این گیرنده است و برای لیگاند سایر GPCR ها اتفاق نمی افتد و به بی حس شدن هومولوگ (همسان) موسوم است.

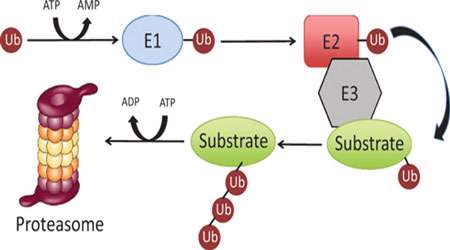
بی حس شدن در RTK ها نیز معمولا با اندوسیتوز (ذخیره) و گاهی تجزیه لیزوزومی گیرنده ها رخ می دهد. اما پیش از آن، غالبا تنظیم کوتاه مدت انجام می گیرد که طی آن، پروتئین فسفاتازها با دومین های اتصال به تیروزین فسفریله (مثل SH2) به گیرنده های فعال فراخوانی می شوند و آنها را دفسفریله و غیر فعال می کنند. این مکانیزم در گیرنده های سایتوکاین هم دیده می شود. در این گیرنده ها، علاوه بر اینها، آنزیم های یوبی کوئتین لیگاز نیز می توانند به گیرنده فراخوانی شوند و JAK ها را برای تجزیه پروتئازومی پلی یوبی کوئتینه کنند.
منبع: http://www.virascience.com http://irisweb.ir
تنظیم کننده: محبوبه همت